
Motion Perception
Contents
Introduction
Motion is defined as a continuous change in an objects location as a function of time. At the level of a single area of the retina, motion is the change in retinal illuminance that occurs at a rate dependent on the speed at which the object moves at its spatial frequency composition.

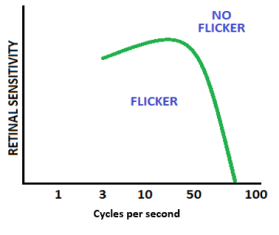
Temporal modulation transfer function
The ability to detect motion is dependent on the temporal resolution of the visual system. The temporal modulation transfer function (temporal CSF), which is seen in the figure to the left, describes temporal resolution (the time required to revisit and aquire detail from the same location on the retina). It has a band-pass function and everything seen below the curve is seen as flicker and everything outside of the curve is not detected as flicker.
The Importance of Motion Detection
Motion detection developed early and is associated with survival, as so predators could spot and hunt their prey and prey could spot and escape predators.
Motion detection allows us to shift attention to the many objects that may be in our periphery and determine 3D shapes through parallax error. It also allows for interaction with the environment and co-ordination of our own movements along with the movements of other moving targets (such as in catching or throwing).
Types of Motion (Top)
There are four types of motion; real, apparent, induced and the motion after-effect. The following subsections will investigate these further.
Real Motion
Real motion is the physical movement of a stimulus against its background (such as a racing car travelling across driving across a scene). This causes a change in luminance across the retina. We also perceive real motion movement when we follow the object with our eyes or our head as so the object remains stationary, but the background does not.
Apparent Motion
Apparent motion is the illusion of motion perceived by looking at a rapid succession of static images which contain sampled real motion of regular intervals. It has the same mechanism as real motion and forms the basis of films, movies, cartoons, television programmes and flipbooks.
In its simplest form, apparent motion is two spatially separated lights turned on and off alternately to perceive smooth motion.
There are two types of apparent motion; beta motion and phi motion.
Beta Motion is the apparent motion where the motion perceived is the same colour as the target. If you alternate the switching on and off of several spatially separated lights at a steady pace, then the motion perceived seems to appear from one location to the other. This can be seen in the figures below:
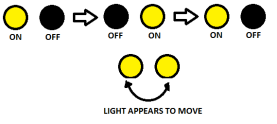

The basics and a demonstration of beta motion
It should be remembered as the illusion of motion where the movement perceived is the same colour and shape as the actual colour of the object/flashing lights. It is beta motion that allows us to create cartoons and motion pictures.
Phi Motion is the motion perceived where there is an alternate switching on and off of lights at a rapid pace. There is no smooth movement detected between the stimuli, but the apparent motion is perceived to be between and around the flickering stimuli. This can be seen in the figures below:
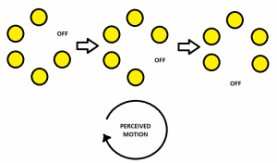
The basics and a demonstration of phi motion
The movement of the apparent motion does not take on any particular shape, but it takes on the colour of the background and not the colour of the targets.
Both beta and phi apparent motion both work is due to persistence of vision - which is where the human perception of the decay of an image is slower than the actual stimulus decay of the image. This means that the original stimulus lingers slightly in the percept and when replaced by the next stimulus in a different position, the image then appears to have moved.
Induced Motion
Induced motion is the movement of one object (usually a larger object) that induces the perception of the movement of another object (usually the smaller object).
An example of this can be seenin the image to the right, demonstrating what happens to our motion perception of the moon on a windy and slightly cloudy night. When the large looking clouds blow across the moon, the direction of the clouds is easy to spot. However, it also appears that the moon is moving in the opposite direction to the clouds, although the moon is not moving in the perceived away at all.
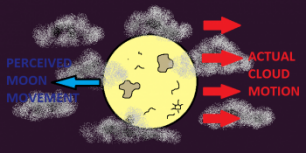
The perceived motion of the moon heading the opposite way to the moving clouds is an example of induced motion
The Motion After-Effect
If a person stares consistently at a movement in a particular direction, then subsequently viewed stationary scenes appear to briefly move in the opposite direction.
This is due to cortical neurone adaptation. The cortical neurones adapt themselves to a given moving stimulus and lower their sensitivity, as to not fire constantly at a high rate for prolonged periods of time. When looking away towards the stationary scene, their reduced sensitivity compared to their neighbouring neurones will cause the perception that movement it occurring in the opposite direction.
This effect is often called the "waterfall effect" as it was first described after looking at the downwards motion of a waterfall followed by the then apparent upwards movement of the cliff when looking away. A similar effect is that called the "belladonna effect", where looking at inward rotating spirals for a period of time will cause the perception of a pupil dilating when subsequently viewed.


The Belladonna Effect: Stare into the centre of the spiral for 30 seconds then look directly into the pupil on the picture of the eye. Notice how the pupil starts to dilate?
The Physiology of Motion Perception (Top)
Motion detection occurs at many points along the visual system. The following subsections will cover the events that happen from retina to cortex.
Retina
At the level of the retina, the basis of motion perception occurs. Temporal resolution is better for cones than rods due to less temporal summation occurring in rods. The magnocellular pathway (parasol cells) have good temporal resolution as opposed to the parvocellular (midget cells) that have poor temporal resolution.
Therefore cone cells in the mid-periphery that utilise the magnocellular pathway contribute most in motion perception. This is because the mid-peripheral retina has the highest density of magnocellular cells, so motion is processed faster on this point on the retina.
The Lateral Geniculate Nucleus (LGN)
The magnocellular retinal ganglion cells project to the magncellular layers of the LGN (layers 1 and 2) and follow the geniculocortical pathway.
However, a small proportion of these retinal fibres project from the retina to the superior colliculus, which aids in eye movement control. These fibres therefore follow the retinotectal pathway.
Area V1
In the visual cortex, there are lots of complex and hypercomplex cells that are direction sensitive. Every person normally has cells sensitive to every direction and representing this on every area of the retina at every possible orientation. The pattern of neurone responses will determine the precise characteristics of the stimulus.
Extra-Striate Cortex
The magnocellular information is further processed in the extra-striate cortex (Area V2) in the thick stripes. Beyond this the information is channelled into the dorsal (where) and the ventral (what) pathways.
The motion information travels in the dorsal (where) pathway to Area V3 and then on to the Middle Temporal (MT) area. 90% of the cells in MT are direction sensitive, with many of these cells also being sensitive to stimulus speed. We know that MT is very important in motion perception as damage to this area can lead to akinetopsia (motion blindness).
From MT, most information travels to the medial superior temporal (MST), which is concerned with optic flow. This essentially determines what is moving in the visual scene when the visual scene itself is moving.
The overall pathway can be seen in the figure below.
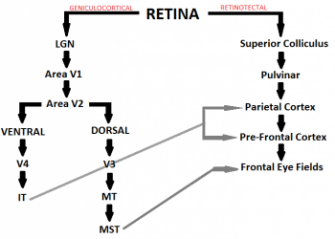
The motion pathway from the retina to the brain
Evidence for Directionally Sensitive Neurones (Top)
Science has come to accept that cortical neurones are directionally sensitive. There are three main pieces of evidence that can support this.
Microstimulation Experiments.
These experiments record the responses from cortical neurones in response to a moving stimulus. It found that cells in both Area V1 and MT are direction selective and the cells in the retina and LGN are not.
Deprivation Experiments.
In 1990, Pasternak reared kittens in a stroboscopic environment. This flashing environment resulted in kittens with vision of still snapshots of their environment. This is due to the elimination of 90% of the directionally sensitive neurones in Area V1. This eliminated the cat's ability to detect direction of movement.
Motion After-Effect. As mentioned earlier, the motion after-effect works due to adaptation of directionally sensitive neurones to a moving stimulus. The effect of this is interocular, meaning that the cells must be located in the brain for the effect to be perceived in the other eye is adapting to the stimulus with one eye only.
With these three pieces of evidence, it is clear that the cells in the cortex are directionally sensitive.
Mechanisms for Encoding Motion (Top)
There are several proposed mechanisms for how motion is encoded in the visual system.
Direction-Sensitive Motion Sensors
This is thought that the motions sensitive cells in the cortex are a lower-level process used to analyse the motion of luminance profiles and short range motion patterns. Short range motion is within the spatial and temporal limits of the receptive fields of the motion detector.
Feature Tracking
Feature tracking is a high level process that locates the position of elements or features in the scene and track their location over time. This is used in long-range motion detection, which is motion that falls outside of the receptive fields of the motion detector (such as beta motion).
Feature tracking requires the ability to match corresponding features and is possibly based on knowledge of heuristics (the knowledge of predicted movements of objects).
THIS CONCLUDES THE UNIT ON MOTION PERCEPTION
BACK TO TOP OF PAGE UNIT 7: BIOLOGICAL MOTION